
Tweet
This week at Infection Landscapes we will cover influenza, one of the most significant respiratory diseases to affect humans, and which does so on an annual basis. Influenza causes substantial morbidity and mortality each year in both the developing and developed world.
The name "influenza" is derived from the Italian word for influence because, centuries ago, it was believed that this disease occurred as a result of the influence of the movement of the sun and the stars given its seasonal nature.
The Pathogen. Influenza is caused by one of three genera of influenza viruses in the Orthomyxoviridae family: influenza A, influenza B, and influenza C. Of these three, influenza A is the most virulent and thus is responsible for the greatest burden of disease. As such, most of this discussion will focus on influenza A.

Influenza A viruses are enveloped and have an RNA genome in 8 segments. The genome is single stranded and negative sense, and is enclosed in a helical capsid:




The Epidemiology and the Landscape. Influenza A virus is spread via droplet, airborne and contact transmission, with droplet transmission probably serving as the most important during seasonal transmission. The overall attack rate in large populations is typically between 10% to 20% during the annual epidemics (seasonal flu), however some subpopulations or local outbreaks can have attack rates approaching 40%. These higher attack rates usually occur in children and, as such, they play a critical role in the spread of influenza in communities during the annual epidemics. Indeed, schools and/or daycare centers will often serve as the primary loci of infection transmission during these epidemics. This is reflected in the typical peak incidence of infection occurring in young school age children prior to the peak in any other age group. The annual global incidence of influenza is unknown given the extreme difficulty in recording and classifying respiratory infections that do not result in hospitalization in most parts of the world. However, the annual global mortality attributable to influenza is estimated to be between 250,000 and 500,000 deaths. In the United States, the annual incidence is estimated to be between 35 and 50 million cases, with between 200,000 and 250,000 hospitalizations and 36,000 deaths.

The above map (published in PLoS Comput Biol 6(1): e1000656. doi:10.1371/journal.pcbi.1000656) simulates the typical distribution of prevalent influenza cases in the US once the seasonal epidemic has begun, given that each infectious case will, on average, lead to more than one new infectious case in a completely susceptible population (R0=1.6). The map demonstrates the high level of transmission, as well as the potential need for the mobilization of resources, in urban areas.
Seasonal, or epidemic, influenza follows a annual, roughly predictable, pattern each year in the temperate regions of the northern and southern hemispheres. In the southern hemisphere, epidemics usually occur in the May to September winter season, while in the northern hemisphere they occur from October to May. For example, in China, Europe, and North America, the epidemic peak(s) usually occurs between November and March: manifested first by high rates of school and industrial absenteeism, followed by an increase in visits to health care facilities, an increase in pneumonia and influenza hospital admissions, and finally an increase in deaths from pneumonia or influenza. In any specific locality, epidemic influenza often begins abruptly, reaches a peak within three weeks, and usually ends by 8 weeks. A given region can also experience 2 epidemic peaks with different virus strains, either in sequence or with overlap. In tropical climates influenza can occur in seasonal epidemics, often during the monsoon or rainy seasons, or endemic transmission can occur throughout the year. The former tropical pattern may suggest that, in these areas of high humidity, powerful climatic forces such as a monsoon may cause people to spend more time indoors in close proximity to each other and thus facilitate spread of disease. Regardless of the time of year, geography, or climate, epidemic influenza occurs because of antigenic drift, i.e. the slow incremental changes that accrue in the viral genome by way of mutation as described above.
Pandemic influenza does not occur in a regular predictable pattern each year, but rather occurs only periodically over the course of decades and can be much more severe because all or a large proportion of the the population is immunologically naive and clinically susceptible to the new emerging virus. These are the antigenic shift events, i.e. widespread global infection following those changes in the viral genome that are due to reassortment. Over the course of the last one hundred years, 5 such shift events have occurred that have lead to 5 subsequent pandemics. The most devastating of these was the 1918 shift to the H1N1 subtype, which resulted in the death of approximately 50 million people worldwide. In 1957, influenza A shifted from the H1N1 to the H2N2 subtype, but with a much lower global burden of disease than the previous 1918 pandemic. In 1968, influenza A H2N2 was replaced by influenza A H3N2, which circulated for the next decade until, in 1977, the influenza A H1N1 subtype re-emerged. This re-emergence was associated with very high attack rates (~50%) in individuals born after 1956 as they had not been exposed because the H1N1 subtype had been replaced by the H2N2 subtype in 1957. For the next three decades, from 1977 until 2009, the re-emerged H1N1 and H3N2 subtypes co-circulated in the population causing the annual influenza epidemics each year. Then, in 2009, a new influenza A H1N1 subtype emerged in Mexico and subsequently replaced the earlier H1N1 subtype. This most recent pandemic in 2009 did not cause the extensive morbidity and mortality that was feared at the beginning of the outbreaks in North America. Overall, morbidity and mortality were lower than in previous pandemics. It is believed that this may have been due in part to the reduced morbidity and mortality among people over the age of 65 years. Nevertheless, there was substantive morbidity and mortality associated with this pandemic and it followed atypical patterns. For example, the vast majority of deaths (~96%) occurred in people under the age of 65, whereas the majority of deaths (~90%) occur in people over the age of 65 during seasonal influenza epidemics. Moreover, the number of deaths due to influenza in children was 4 times greater during the 2009 pandemic than what it is during seasonal influenza epidemics.
It is important to discuss the ecologic relationship between human epidemic influenza and pandemic influenza. To begin, there are strains of influenza A virus that infect humans, there are strains that infect birds, and there are strains that infect pigs. Nevertheless, all influenza A viruses circulating today are evolutionarily derived from bird influenza A. Birds, and in particular waterfowl, are the natural primary reservoir host for all current circulating bird inlfuenza A. These birds are also, either directly or indirectly via another intermediate host (generally pigs), the historic reservoir for all influenza A that subsequently adapted to other animals. New subtypes of human influenza A typically emerge when the genome of a current circulating human influenza A subtype reassorts with the genome of a subtype of current circulating bird influenza A or the genome of a subtype of current circulating pig influenza A. Pigs can harbor infection with several influenza subtypes because they have the epithelial cell receptors for both bird and human HA. As such, pigs are considered important "mixing vessel" hosts for reassortment events. However, in order for a new human subtype to become a pandemic virus it must 1) be new to a relatively large proportion of the population; i.e. the global population must be relatively immunologically naive to the new subtype, 2) it must be able to replicate and cause disease in humans, and 3) it must be easily transmissible between humans. The last point is particularly important as the new subtype will not become a human inlfuenza A subtype if it cannot be transmitted between people, even if human infection is possible zoonotically. Infection with the highly pathogenic avian influenza A H5N1 (commonly referred to as bird flu) exemplifies this. This is a bird inlfuenza A subtype, not a human inlfuenza A subtype. While this bird subtype is capable of infecting and causing severe disease in some humans, it is not currently capable of causing a pandemic because it cannot be transmitted efficiently from person to person. The graphic below produced by the National Institute of Allergy and Infectious Diseases nicely depicts the species and transmission pathways that can lead to antigenic shift in the influenza A virus and subsequent pandemics.

The mixing of different influenza A subtypes from different animal species is central to producing new and potentially dangerous subtypes that can result in new human influenza A, which can subsequently lead to new pandemics. Moreover, the nature of such genetic exchange makes the elimination of human influenza A virtually impossible. There are two primary landscapes that converge to optimize reassortment of influenza A subtypes: 1) the sylvan landscapes and migratory paths of waterfowl and, 2) the agricultural lands and markets, both subsistence and industrial, of humans. The spaces where these converge are places where sylvan birds can come into contact with domestic birds (e.g. chickens), which may also be in direct contact with pigs. As such, these landscapes can be important sources of diverse influenza A subtype circulation among multiple hosts and may serve as important sources of human exposure.

This week at Infection Landscapes we will cover influenza, one of the most significant respiratory diseases to affect humans, and which does so on an annual basis. Influenza causes substantial morbidity and mortality each year in both the developing and developed world.
The name "influenza" is derived from the Italian word for influence because, centuries ago, it was believed that this disease occurred as a result of the influence of the movement of the sun and the stars given its seasonal nature.
The Pathogen. Influenza is caused by one of three genera of influenza viruses in the Orthomyxoviridae family: influenza A, influenza B, and influenza C. Of these three, influenza A is the most virulent and thus is responsible for the greatest burden of disease. As such, most of this discussion will focus on influenza A.

Influenza A viruses are enveloped and have an RNA genome in 8 segments. The genome is single stranded and negative sense, and is enclosed in a helical capsid:

Hemagglutinin (HA) and neuraminidase (NA) are important surface glycoproteins that facilitate virion entry into and exit from the host cell. As such, these surface structures are key features of virus infectivity and pathogenicity. HA functions as the attachment glycoprotein to the cellular receptor, sialic acid, on columnar epithelial cells of the respiratory tract. Thus, HA specificity for receptor binding is a determinant of which species can be infected, i.e. the host range, which will be discussed in more detail later. Specific antibodies to HA epitopes prevent attachment and entry of the influenza virus particle into the host cells, and are thus an important aspect of immune protection against infection. NA cleaves sialic acid residues to allow virus particle release from the host epithelial cell. It is suspected that specific anti-NA antibodies diminish the release of new virions from host cells, thus curtailing an infection. The infection cycle of the virus in the host cell is very nicely depicted in the graph below published by Influenza Report:

The animated video below also provides a nice depiction of the infectivity, pathogenicity, and immunogenicity of influenza A.
Antigenic variation is very important to influenza A evolution, particularly as this relates to the annual epidemics and multidecadal pandemics experienced by humans. Before talking more specifically about these two phenomena, it will be helpful to highlight two features of the molecular biology of the virus that are particularly important to its rate of genetic change. First, the genome is comprised of RNA molecules. Many such RNA viruses, including influenza A, exhibit low fidelity and, thus, a high rate of mutation because they lack the proofreading provided by DNA polymerase, which is a necessary enzyme for DNA virus assembly. Second, the genome of influenza A virus is segmented, which further contributes to the high rate of genomic change because the genes encoded on particular segments of one genome can more easily be exchanged with segments of other influenza A genomes from multiple virus particles that happen to infect the same host cell.
There are two fundamental evolutionary genetic processes at play in the antigenic properties of influenza A virus. The first is antigenic drift. Antigenic drift defines the minor yet frequent genetic changes in the HA and NA surface antigens that occur on an annual or near-annual basis. Antigenic drift leads to approximately 1% change in the HA antigen per year. This rate of mutation is the leading force driving the annual epidemics experienced every year around the world during the regular influenza season. However, there is a second, equally important, process involved. The second is antigenic shift. Major changes in the amino acid composition for the HA or NA surface antigens, or both, can lead to a new viral subtype. These new subtypes will present (in most instances) to a completely immunologically naive population wherein everyone, or nearly everyone, is susceptible to infection. It is believed that after several decades (typically between 10 and 30 years) of circulation of a specific influenza A subtype, most members of the population will have antibody to that subtype. This accumulated population immunity places selective pressure on the virus and leads to new shift variants that emerge by way of reassortment. The new subtypes lead to pandemic influenza events.

The patterns of transmission during the annual epidemics that correspond to antigenic drift, and the multidecadal pandemics that correspond to antigenic shift will be described later in the epidemiology section.
The Disease. Clinical disease typically presents with abrupt onset of fever and respiratory symptoms, including cough, rhinorrhea and sore throat. Malaise and prostration are very common. Myalgia and headache are more common with influenza than with other respiratory infections. In addition, gastrointestinal symptoms are not common in adults, but 50% of infant and child infections may have vomiting, abdominal pain and diarrhea. However, none of these symptoms are definitive for influenza infection. Only cell culture demonstrating influenza A virus can distinguish influenza from other agents causing respiratory infection, such as a rhinovirus (a frequent source of the "common cold"). Nevertheless, influenza infection is on average associated with more severe disease than the pathogens responsible for the common cold. Symptomatic disease usually lasts 3 to 5 days, but complications can prolong the illness.

The complications associated with influenza make the infection particularly dangerous, especially for infants, the elderly, and individuals who are immunocompromised. Some individuals can develop primary viral pneumonia, which frequently can be fatal. However, primary viral pneumonia is not very common. Secondary bacterial pneumonia is much more common, particularly among the susceptible groups mentioned above, and can occur up to two weeks after the acute viral infection. In infants and children, otitis media and croup are common complications. Myocarditis, myositis and encephalitis are less frequent complications.
The Epidemiology and the Landscape. Influenza A virus is spread via droplet, airborne and contact transmission, with droplet transmission probably serving as the most important during seasonal transmission. The overall attack rate in large populations is typically between 10% to 20% during the annual epidemics (seasonal flu), however some subpopulations or local outbreaks can have attack rates approaching 40%. These higher attack rates usually occur in children and, as such, they play a critical role in the spread of influenza in communities during the annual epidemics. Indeed, schools and/or daycare centers will often serve as the primary loci of infection transmission during these epidemics. This is reflected in the typical peak incidence of infection occurring in young school age children prior to the peak in any other age group. The annual global incidence of influenza is unknown given the extreme difficulty in recording and classifying respiratory infections that do not result in hospitalization in most parts of the world. However, the annual global mortality attributable to influenza is estimated to be between 250,000 and 500,000 deaths. In the United States, the annual incidence is estimated to be between 35 and 50 million cases, with between 200,000 and 250,000 hospitalizations and 36,000 deaths.

The prevalence of influenza in a single simulation of the United States 100 days after the start of an influenza epidemic with R0=1.6. The color of each dot corresponds to the illness prevalence in a census tract.
The above map (published in PLoS Comput Biol 6(1): e1000656. doi:10.1371/journal.pcbi.1000656) simulates the typical distribution of prevalent influenza cases in the US once the seasonal epidemic has begun, given that each infectious case will, on average, lead to more than one new infectious case in a completely susceptible population (R0=1.6). The map demonstrates the high level of transmission, as well as the potential need for the mobilization of resources, in urban areas.
Seasonal, or epidemic, influenza follows a annual, roughly predictable, pattern each year in the temperate regions of the northern and southern hemispheres. In the southern hemisphere, epidemics usually occur in the May to September winter season, while in the northern hemisphere they occur from October to May. For example, in China, Europe, and North America, the epidemic peak(s) usually occurs between November and March: manifested first by high rates of school and industrial absenteeism, followed by an increase in visits to health care facilities, an increase in pneumonia and influenza hospital admissions, and finally an increase in deaths from pneumonia or influenza. In any specific locality, epidemic influenza often begins abruptly, reaches a peak within three weeks, and usually ends by 8 weeks. A given region can also experience 2 epidemic peaks with different virus strains, either in sequence or with overlap. In tropical climates influenza can occur in seasonal epidemics, often during the monsoon or rainy seasons, or endemic transmission can occur throughout the year. The former tropical pattern may suggest that, in these areas of high humidity, powerful climatic forces such as a monsoon may cause people to spend more time indoors in close proximity to each other and thus facilitate spread of disease. Regardless of the time of year, geography, or climate, epidemic influenza occurs because of antigenic drift, i.e. the slow incremental changes that accrue in the viral genome by way of mutation as described above.
Pandemic influenza does not occur in a regular predictable pattern each year, but rather occurs only periodically over the course of decades and can be much more severe because all or a large proportion of the the population is immunologically naive and clinically susceptible to the new emerging virus. These are the antigenic shift events, i.e. widespread global infection following those changes in the viral genome that are due to reassortment. Over the course of the last one hundred years, 5 such shift events have occurred that have lead to 5 subsequent pandemics. The most devastating of these was the 1918 shift to the H1N1 subtype, which resulted in the death of approximately 50 million people worldwide. In 1957, influenza A shifted from the H1N1 to the H2N2 subtype, but with a much lower global burden of disease than the previous 1918 pandemic. In 1968, influenza A H2N2 was replaced by influenza A H3N2, which circulated for the next decade until, in 1977, the influenza A H1N1 subtype re-emerged. This re-emergence was associated with very high attack rates (~50%) in individuals born after 1956 as they had not been exposed because the H1N1 subtype had been replaced by the H2N2 subtype in 1957. For the next three decades, from 1977 until 2009, the re-emerged H1N1 and H3N2 subtypes co-circulated in the population causing the annual influenza epidemics each year. Then, in 2009, a new influenza A H1N1 subtype emerged in Mexico and subsequently replaced the earlier H1N1 subtype. This most recent pandemic in 2009 did not cause the extensive morbidity and mortality that was feared at the beginning of the outbreaks in North America. Overall, morbidity and mortality were lower than in previous pandemics. It is believed that this may have been due in part to the reduced morbidity and mortality among people over the age of 65 years. Nevertheless, there was substantive morbidity and mortality associated with this pandemic and it followed atypical patterns. For example, the vast majority of deaths (~96%) occurred in people under the age of 65, whereas the majority of deaths (~90%) occur in people over the age of 65 during seasonal influenza epidemics. Moreover, the number of deaths due to influenza in children was 4 times greater during the 2009 pandemic than what it is during seasonal influenza epidemics.
It is important to discuss the ecologic relationship between human epidemic influenza and pandemic influenza. To begin, there are strains of influenza A virus that infect humans, there are strains that infect birds, and there are strains that infect pigs. Nevertheless, all influenza A viruses circulating today are evolutionarily derived from bird influenza A. Birds, and in particular waterfowl, are the natural primary reservoir host for all current circulating bird inlfuenza A. These birds are also, either directly or indirectly via another intermediate host (generally pigs), the historic reservoir for all influenza A that subsequently adapted to other animals. New subtypes of human influenza A typically emerge when the genome of a current circulating human influenza A subtype reassorts with the genome of a subtype of current circulating bird influenza A or the genome of a subtype of current circulating pig influenza A. Pigs can harbor infection with several influenza subtypes because they have the epithelial cell receptors for both bird and human HA. As such, pigs are considered important "mixing vessel" hosts for reassortment events. However, in order for a new human subtype to become a pandemic virus it must 1) be new to a relatively large proportion of the population; i.e. the global population must be relatively immunologically naive to the new subtype, 2) it must be able to replicate and cause disease in humans, and 3) it must be easily transmissible between humans. The last point is particularly important as the new subtype will not become a human inlfuenza A subtype if it cannot be transmitted between people, even if human infection is possible zoonotically. Infection with the highly pathogenic avian influenza A H5N1 (commonly referred to as bird flu) exemplifies this. This is a bird inlfuenza A subtype, not a human inlfuenza A subtype. While this bird subtype is capable of infecting and causing severe disease in some humans, it is not currently capable of causing a pandemic because it cannot be transmitted efficiently from person to person. The graphic below produced by the National Institute of Allergy and Infectious Diseases nicely depicts the species and transmission pathways that can lead to antigenic shift in the influenza A virus and subsequent pandemics.
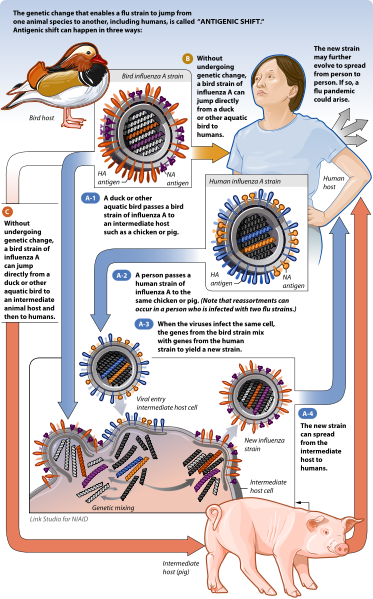
The mixing of different influenza A subtypes from different animal species is central to producing new and potentially dangerous subtypes that can result in new human influenza A, which can subsequently lead to new pandemics. Moreover, the nature of such genetic exchange makes the elimination of human influenza A virtually impossible. There are two primary landscapes that converge to optimize reassortment of influenza A subtypes: 1) the sylvan landscapes and migratory paths of waterfowl and, 2) the agricultural lands and markets, both subsistence and industrial, of humans. The spaces where these converge are places where sylvan birds can come into contact with domestic birds (e.g. chickens), which may also be in direct contact with pigs. As such, these landscapes can be important sources of diverse influenza A subtype circulation among multiple hosts and may serve as important sources of human exposure.

Control and Prevention. There are four important tactics that have demonstrated effectiveness in either preventing infection or mitigating the effects of an active infection. The first is very simple: regular hand washing during the flu season. This certainly need not be obsessive, but the regular practice of washing your hands with regular bar soap (NOT antibacterial soaps or hand sanitizers, which can be harmful to our microbiome) after returning home from populated public spaces during flu season makes good sense. The second is even more simple, though rarely followed: stay home when sick. The likelihood of transmitting influenza virus to susceptible people is quite high during illness. In addition, given that influenza virus is typically more virulent than many of the respiratory pathogens associated with the common cold, an infected individual requires significant rest to convalesce. As such, going in to work or school during a respiratory illness can be dangerous both to oneself and one's peers. Third, the use of antiviral drugs can be used to reduce the duration of symptoms among infected individuals and may help prevent transmission to their susceptible contacts. There are two classes of these drugs: the neuraminidase inhibitors, oseltamivir and zanamivir, and the M2 inhibitors, amantadine and rimantadine. The neuraminidase inhibitors block the release of new virions from the surface of infected host cells. The M2 inhibitors block the release of viral RNA into the host cell after the virion has entered the cell. The fourth important measure of control and prevention is vaccination.
The previous discussion of antigenic drift highlighted the importance of influenza virus mutability. This requires that a new vaccine is produced each year because the vaccines stimulate an immune response that targets those same components of HA and NA antigens whose genes are subject to the high rate of mutation. The process of having to renew vaccine production with new strains of virus on an annual basis has made achieving a consistently high level of vaccine effectiveness difficult. Nevertheless, it is currently the best we have and the vaccine effectiveness is not negligible.
There are two types of vaccine: an inactivated vaccine (the injection) and a "live" attenuated vaccine (nasal spray). Neither of these vaccines are capable of causing influenza. The overall vaccine effectiveness for the inactivated vaccine against seasonal influenza based on recent meta-analyses is approximately 60% (the "live" vaccine seems to have a higher effectiveness, however the data are much more limited because so much less of the this vaccine is produced). While this is certainly not ideal, it still does provide a level of protection that can have some important implications. First, specific subpopulations, such as school children, can be very important for targeted vaccination as these are typically the loci of epidemic transmission and community dissemination of seasonal influenza. Moreover, there is evidence that the vaccine effectiveness is markedly better in children. Blocking transmission in this setting, even with imperfect effectiveness, can have important implications for the epidemic curves of localized outbreaks, as well as the national incidence of severe illness and hospitalizations and deaths. Second, the vaccine, even when it does not prevent infection for a given individual, can still mitigate the severity of that infection. As such, the people who are vaccinated and do get infected still benefit from protection because they will generally experience illness that is less severe than those who were not vaccinated. And, as discussed above, the problem with influenza is the potential for developing complications when illness is severe. Third, there are subgroups in the population who are at particularly high risk for influenza infection. These are infants and small children, the elderly, and immunocompromised people. By getting vaccinated, we significantly reduce the risk of transmitting what could be a deadly infection to these high risk people by either preventing the infection in ourselves, or by limiting the severity of disease we experience if we do get infected and thus our transmissibility to others. If you are wondering whether or not the influenza vaccine is for you just read through the infographic below by TeleVox:
The previous discussion of antigenic drift highlighted the importance of influenza virus mutability. This requires that a new vaccine is produced each year because the vaccines stimulate an immune response that targets those same components of HA and NA antigens whose genes are subject to the high rate of mutation. The process of having to renew vaccine production with new strains of virus on an annual basis has made achieving a consistently high level of vaccine effectiveness difficult. Nevertheless, it is currently the best we have and the vaccine effectiveness is not negligible.
There are two types of vaccine: an inactivated vaccine (the injection) and a "live" attenuated vaccine (nasal spray). Neither of these vaccines are capable of causing influenza. The overall vaccine effectiveness for the inactivated vaccine against seasonal influenza based on recent meta-analyses is approximately 60% (the "live" vaccine seems to have a higher effectiveness, however the data are much more limited because so much less of the this vaccine is produced). While this is certainly not ideal, it still does provide a level of protection that can have some important implications. First, specific subpopulations, such as school children, can be very important for targeted vaccination as these are typically the loci of epidemic transmission and community dissemination of seasonal influenza. Moreover, there is evidence that the vaccine effectiveness is markedly better in children. Blocking transmission in this setting, even with imperfect effectiveness, can have important implications for the epidemic curves of localized outbreaks, as well as the national incidence of severe illness and hospitalizations and deaths. Second, the vaccine, even when it does not prevent infection for a given individual, can still mitigate the severity of that infection. As such, the people who are vaccinated and do get infected still benefit from protection because they will generally experience illness that is less severe than those who were not vaccinated. And, as discussed above, the problem with influenza is the potential for developing complications when illness is severe. Third, there are subgroups in the population who are at particularly high risk for influenza infection. These are infants and small children, the elderly, and immunocompromised people. By getting vaccinated, we significantly reduce the risk of transmitting what could be a deadly infection to these high risk people by either preventing the infection in ourselves, or by limiting the severity of disease we experience if we do get infected and thus our transmissibility to others. If you are wondering whether or not the influenza vaccine is for you just read through the infographic below by TeleVox:
